


Robert L. Kelly, The Behavioral Ecology of Foragers
Methodological individualism and foragers, including optimal patron clientelism

Robert L. Kelly wrote:
This chapter reviews the use of human behavioral ecology (HBE) in the social sciences [and its application] in the study of hunter-gatherers, or foragers …
… HBE employs two assumptions: methodological individualism and optimization. Selection operates on variability and favors individuals whose behavior enhances the opportunity to increase fitness. Thus, individual choices rather than cultural norms are the focus. However, HBE recognizes that an individual’s goals come from biological and cultural information—cultural norms. The drive to “succeed” entails not only biological directives (to reproduce) but cultural norms as well (e.g., bring home as much meat as possible, spend time with offspring, acquire prestige or wealth). Goals must be defined, but HBE does not assume that the goals of all members of a society will be identical. HBE requires only that individuals be capable of storing knowledge and of understanding (or at least thinking that they understand) the relationship between their actions and goals. …
… Although we can find societies where individuals consciously strive to maximize foraging efficiency or prestige, we do not find societies that consciously value “reproductive fitness”. Behavioral ecologists argue that individuals who meet culturally defined standards of success should manifest a behavioral variant that achieves greater fitness than other variants. …
… HBE claims that the biological capacity for culture must have arisen through a process of natural selection. Individuals who were cultural beings, at some point in the past, became more prevalent in a population at the expense of those hominins who were not cultural. For some this means that cultural behavior is an extension of biological adaptation and operates for similar purposes. Indeed, humans pass on information between generations about kinship, subsistence, religion, morals, aesthetics, and so on. …Recall that evolution is the differential persistence of information—some of that information is genetic, but for humans, much of it is cultural. Cultural information is passed on through enculturation rather than reproduction.
We receive our genes from our biological parents, but we acquire our culture from many people.
Many anthropologists argue that diversity in human behavior is the manifestation of various cultural values that can only be understood within particular cultural and historical contexts. However, in any given culture at any given time, each individual represents slight variations of cultural norms, and culture change is change in the frequency of these variants. …
… In the remainder of this chapter, we consider those areas of life in foraging societies where HBE has been applied: foraging, mobility, technology, demography, food sharing, territoriality, and social and political organization. The beauty of HBE is that in all these cases, the simple models used can accommodate the many specific variables of human life and lead to predictions for realms of behavior not initially covered by the models; it is a remarkably productive paradigm. …
Foraging Behavior:
… An important aspect of optimal foraging models is … marginal value theorem (MVT). The original intent of this theorem was to predict when an animal should leave one foraging patch for another. The MVT models common sense: a forager enters a patch, starts collecting berries, and experiences a high return rate. As the patch is depleted, berries become harder to find, and the overall foraging return rate declines. If the forager remains long enough, searching for the last berry, he or she will have to eat the berries already gathered—and the return rate will become negative. The MVT seeks the rule to use in order to know when to leave the patch for another. …
… The animal foragers for whom foraging models were initially developed tend to be feed-as-you-go, but humans are central place foragers—they harvest food in the field and then return to camp with it. To do so, humans process the food partially—butcher an animal to some degree, or remove pinyon pine seeds from their cones. But time spent processing is time not spent harvesting. The MVT helps because the question “how much should we process a resource in the field?” is similar to “how long should we forage in this patch?” And again, it has to do with distance: process the resource until the return rate, taking travel time to camp into consideration, is equivalent for the unprocessed and processed resource.
Mobility:
Most hunter-gatherers are nomadic, moving in response to the availability of food. We can anticipate how frequently they move with the MVT. As foragers deplete a camp’s immediate area, they must travel farther. As they do, they devote more time and energy to walking, and less to foraging. At some point those foragers will decide it is worthwhile to forgo food collection and invest time in moving camp to a new location. Ethnographic data suggest that the maximum daily foraging radius from camp is about 10 km (beyond this, foragers will remain away from camp at night). As expected from the MVT, foragers often move after exploiting food within a <5 km radius. Many variables affect this relationship and the specific outcome: distance to the next camp, the terrain’s difficulty, the time it takes to manufacture housing, and travel technology.
This perspective on mobility sheds a different light on sedentism—the lack of camp mobility. And this matters because sedentary communities are associated with nonegalitarian sociopolitical organization. Why stop moving? One might assume that the year-round availability of food within a reasonable foraging radius is a necessary condition for hunter-gatherers to become sedentary and remain hunter-gatherers rather than become agriculturalists. But is it a sufficient condition? The MVT predicts that even if a location could be occupied for a year, it would be abandoned long before as a function of diminishing returns. In fact, the MVT suggests that the only reason hunter-gatherers would not move is if there is no place to move to. And the most likely impediment would be high population density and groups inhabiting every habitable place on the landscape. Such a situation could encourage sedentism since the alternative would require the cost of physically displacing another group.
This implies that sedentary villages are associated with control of a resource extraction point and that sedentism has a domino effect: when one group becomes sedentary, it increases the cost of moving for everyone else. As in the game of musical chairs, other groups will grab their own places on the landscape—and through a particular historical process. The ideal-free distribution points out that once the returns from the best area have declined to the point where they are equal to the highest rate from a neighboring, less productive area, some people will move to that less productive area. This predicts that in a region undergoing population pressure, over time specific locales will be occupied by sedentary groups in their rank order of foraging returns; some archaeological case studies support this. …
Territoriality:
The economic-defensibility model (EDM) focuses on the cost and benefit of defending resources. Similar to tolerated theft [MGH: discussed earlier, we skipped it], it argues that territoriality occurs when the cost of defending a resource is less than the benefit derived from it. If a resource is not very dense and its occurrence in time and space is unpredictable, then the cost of defending it could be so high as to offset any gains from its exclusive use. Where resources are dense and predictable, they are worth the effort of defense and exclusive use. This model accounts for some of the variability we see among foragers’ territorial behavior.
Dense, predictable resources can be defended at low cost, because only a small area must be defended. But with population growth, demand will increase and at some point defense will require increased investment, in perimeter defense (building walls, patrolling borders, attacking trespassers, and making retaliatory or preemptive raids). When a population reaches carrying capacity, the benefit of trying to take a resource becomes worth the potential cost and, consequently, so too does defending a resource. What happens, though, where resources are neither dense nor predictable?
Perimeter defense is rare among foragers; more commonly, they acquire permission to use land that “belongs” to another group, and that permission is virtually ensured. Resources are not there for the taking, but they may be there for the asking. The giving of permission is the giving of a gift—and is similar to sharing. And as with sharing, there are costs and benefits to allowing others in: the visitors may reduce the host’s foraging efficiency, but they also must reciprocate in the future. … [It has been] argued that the benefits outweigh the costs when resources are scarce and (consequently) territories are large, making them difficult to patrol, and when food productivity is variable across the landscape, meaning that the tables may be turned in a future year. Under these conditions, foragers ensure access to the resources of another group by maintaining social access to that group, and they can do this through myriad social conventions. Trespass is possible but is a poor decision because trespassers may not have sufficient knowledge of a region to make use of it, and if trespassers are detected (through their tracks, smoke from fires, etc.) they risk retaliation and future exclusion.
We can model the conditions for perimeter versus social boundary defense by modifying [a] model of food sharing. Here we use variance in a residential group’s return rate over time and the degree of correlation in returns of different groups. High variance means that in some years a group has plenty of food and in other years experiences disaster; low variance means that the group always collects about the same amount of food. High correlation means that when one group is doing well, the other is also doing well, and when one group is doing poorly, so is the other; low correlation means that when one group is doing poorly, another is doing well. …
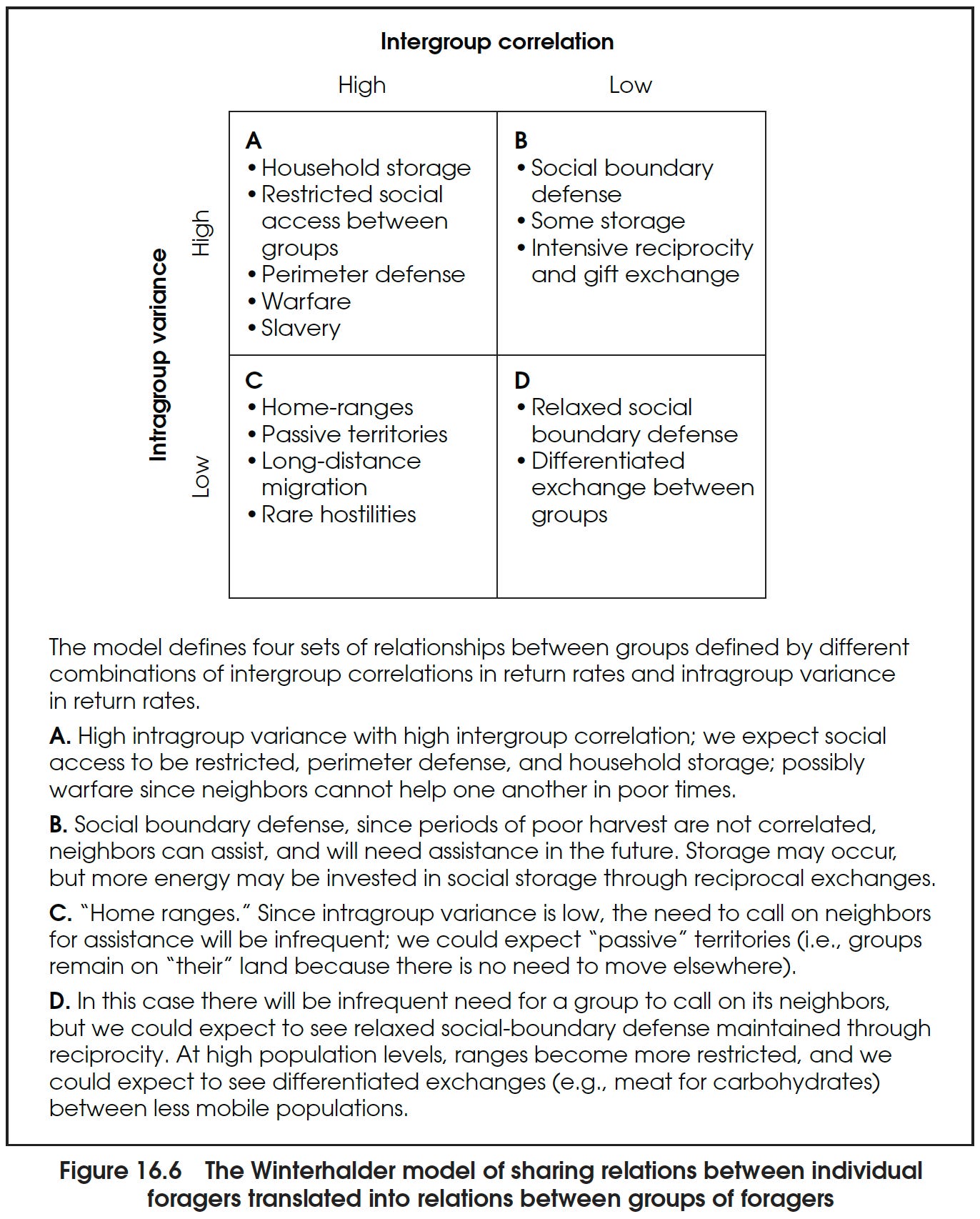
Group Size:
Nomadic foraging societies live in small groups of eighteen to thirty persons [references]. But foragers periodically gather into larger groups, for winter camps, communal fishing, and so on. [Some researchers] found that these various groups tend to increase by a regular factor of four: families consist of 4 or 5 people; a residential group of about 14–17; social aggregations, for example, at winter camps, of 50–60; periodic aggregations of 150–180; and entire “ethnic” populations of 730–950. They argue that these groupings reflect a subliminal consensus about how to most efficiently move food, information, and mates among foragers. For example, consisting of about twenty-five persons, residential group size balances the desire to minimize variance in food intake (through sharing) with the desire to minimize the rate of food depletion (which reduces the frequency of movements), maximizes group size without the need for a decision-making hierarchy, and maintains reproductive viability.
Of these, daily foraging needs are perhaps the most immediate factor in determining group size. [One researcher] pointed out that per capita foraging return rates can increase with group size—up to a point, after which additional group members do not increase work efficiency but, by taking a share, reduce the per capita return rate. This sets up conflict once the optimal group size is reached, since some members will still wish to join if their return rate will improve over what they would achieve by foraging alone. Using data on Inujjuamiut hunting parties, [one researcher] found that groups were slightly larger than the model anticipated, and suggested that this reflected a compromise between a goal of maximizing foraging efficiency and minimizing the risk of social discord.
This may explain why [one] study found that residential groups are frequently larger than expected by the 4× factor of increase. It’s possible that the optimal co-residing foraging group size for nomadic foragers in many different environments is rather small—fifteen people or so—but that long-term social needs and the avoidance of retaliation results in slightly larger residential groupings. Crucial to studies of group size, territoriality, sharing—everything, in fact—is population growth. A few decades ago researchers assumed that foragers consciously kept their populations in check through contraception, abortion, postpartum taboos on intercourse, and infanticide … suffice it to say that there is no evidence these methods work, or that infanticide, outside of some Arctic societies, has a significant impact on population growth. Although hunter-gatherers have long spacing between births (three to four years), there is no evidence that this is intentional, but is instead a product of women’s reproductive ecology. …
… Finally, lethal violence—homicide and warfare—can also reduce population growth rates. However, the rates of both seem to vary with population pressure and consequently the cost-benefit ratio of fighting—as population pressure on food resources increases, so too does the incidence of warfare and homicide. Warfare, organized battles between two groups, is more prevalent among sedentary than among nomadic hunter-gatherers. While there is violence among nomadic foragers, this tends to be of the sort that occurs in egalitarian societies where tensions may eventually erupt into passionate fisticuffs and occasionally lethal violence.

Division of Labor:
A strong empirical and universal pattern among foragers is that men hunt large game and women gather plant foods and small game. This has been a “political” issue, and rightfully so: hunting requires skill and patience, not extraordinary strength or intelligence. But since foraging children breastfeed until they are several years old (due to the lack of weaning foods), children remain with their mothers. Children reduce a woman’s efficiency when foraging for sessile plant foods, and they would greatly reduce a woman’s hunting efficiency. Women do not hunt, simply because, with children, hunting large game would produce a lower return rate than plant-food collection. Women undertake child-compatible hunting (e.g., communal net hunting) when that provides higher returns than other foraging opportunities. The importance of this observation is that men are the ones who “own” the highly desirable and sharable resource of large game.
Social Organization:
There has been relatively little HBE work into elements of social organization such as kinship, descent, postmarital residence, and marriage (with the exception of polygyny). However, if we look at these elements as manifestations of strategies of social alignment, they can be related to the group size model discussed above. For example, long-distance large-game hunting or warfare removes men from their families for long periods of time and places them at risk. This seems to result in matrilocal postmarital residence as part of a woman’s reproductive strategy to ensure that she has kin nearby to assist in child rearing. Rules of residence are related to a number of variables, but all revolve around individuals finding ways to join one group or another. For men, the primary factors may be foraging or warfare, whereas for women it may be assistance in child rearing. …
Political Organization:
Hunter-gatherers are known for their egalitarian political organization, which is marked primarily by autonomy, more or less individual control over their lives. But some foragers are distinctly nonegalitarian, and the historical shift that brings this change about is complex. It seems to occur only where foragers are sedentary or nearly so, where a key resource-extraction locality can be controlled through territorial behavior, and where population pressure is high. In this context, selection works against egalitarianism in favor of nonegalitarian behaviors.
The patron-client model [references] describes the political outcome of sedentary foragers. We can give only an outline of the entire process here. Imagine a group of sedentary foragers living in a region of high population pressure who are situated on a productive resource that fluctuates little from year to year. … [T]his group has little incentive to share with other, less fortunate groups in the region because the sedentary foragers do not anticipate needing assistance. However, the less fortunate foragers have every reason to try (especially in bad years) to gain access to what the well-positioned group has. They are a constant threat, and one village might buy off another. Recalling the group-size argument, members beyond the “optimal” number might be admitted, through marriage or kinship connections, but at a price: secondary status, which ideologically justifies a reduced share even when contributing actual work. At that moment, nonegalitarian society forms.
This can happen especially where the per capita return rate increases with a larger group size … [and] the benefits of specialization … Leaders arise in such situations if they take on the cost of ejecting the free-riders who can invade large groups and the cost of coordinating efforts to avoid inefficiency. Their importance is relative to the difference between foraging alone and as part of a group, to the size of the optimal group, and to the extent that a resource-extraction locale is defensible.
In return, the leader takes a slightly larger cut of the foraging returns. Hard-working group members benefit because their efforts are not supporting goldbrickers, and yet they do not have to pay the social cost of “being stingy”. In economies of scale, foragers benefit in the long term by turning over some of their autonomy to a leader and reducing their maximum possible return rate.
Assuming a leader skims … the group’s average per capita utility curve and the utility curve of the leader are similar to that of Figure 16.7 [above], but that the utility curve of the leader predicts a slightly larger group size. It is to a group leader’s advantage to keep members in the group, even when it is not necessarily to the other members’ maximum benefit. Culture assists the process, for a leader does not have to maintain a position through brute force, but through the force of ideology, belief systems that “explain” inequality. …
This ideology can entail warfare, costly displays, and “generosity”. Warfare acquires more resources for loyal followers and preempts attacks. In large groups, a smaller percentage of people are needed for warfare, and thus some can avoid defense’s direct cost, free-riding on the efforts of others. Costly displays, such as expensive public feasts, demonstrate a leader’s power to would-be adversaries by being an honest signal of the numerical strength of a population and the capacity of a leader to mobilize that strength.
Finally, generosity (sometimes through the spoils of war) can buy off the competition and solidify the patron-client relationship. Through a process similar to tolerated theft, additional increments of wealth have less value with increasing amounts of overall wealth. The same increase in wealth holds different potential benefits. Currently wealthy individuals can buy off less wealthy ones because an increase of X amount for them is simply not worth as much as it is to the less wealthy. At high levels of wealth, the social benefits of generosity are worth more than the resource itself. Through this process, inequality tends to propagate itself until some process … literally overthrows the situation.
Conclusion:
… HBE offers a productive research strategy for understanding the mechanism linking environment and society. It offers a perspective and methodology that help us understand how a forager … makes decisions. And it does so with a firm grounding in evolutionary theory. Behavioral ecology begins with the assumption that people’s behavior is aimed at maximizing reproductive success. This is a provisional assumption; culture can alter the process, changing what is meant by “maximize”.
I have focused on foragers in this chapter because that is my specialty, but also to make a point. Foragers play an especially important role in evolutionary research paradigms, because, as the argument goes, our capacities were created in the Pleistocene, when all humans lived by hunting and gathering. Many researchers, especially those outside anthropology, assume that in looking at living foragers they are looking back into the past. This is a dangerous supposition. Living foragers do live under circumstances that are closer to those of our most ancient ancestors than to those of, say, the average university professor. But living foragers are also affected by their social context, which today includes World Bank–sponsored projects, terrorist movements, government-sponsored resettlement programs, tourism, and market craft production. HBE does not assume and does not have to assume that living foragers are pristine relics of the Paleolithic. Living foragers, even those who use guns or snowmobiles or who work at wage labor, provide us with hypotheses and tests of models; but when we look at the modern Hadza, Ache, Efe, or Ju/’hoansi, we are not looking back in time. Too many researchers today make this mistake. Likewise, too many researchers make claims about how foraging lifeways created the human mind (e.g., male-female differences in spatial abilities) when they in fact know little about those lifeways. Hopefully, this chapter can help correct that problem.
The Source:
Robert L. Kelly, ‘The Human Behavioral Ecology of Foragers’ in Handbook on Evolution and Society : Toward an Evolutionary Social Science, edited by Jonathan H. Turner, Richard Machalek, Alexandra Maryanski, Routledge 2015
Evolutions of social order from the earliest humans to the present day and future machine age.